Patogenia del virus de la gripe porcina y su relación con la aparición de complicaciones bacterianas secundarias
28 abril, 2021Artículo publicado en Suis nº 167 - Mayo 2020
Gerard Martín
Investigador del CReSA
Introducción
La dinámica de infección del virus de la gripe A (VIA) en explotaciones es compleja. Existe en dos formas principalmente; la forma epizoótica, en la que se observa una gran incidencia de la infección en prácticamente la totalidad de la población expuesta; y la forma enzoótica, en la que la infección ocurre de forma recurrente lote tras lote productivo con una incidencia variable y con un impacto clínico difícil de medir. En esta segunda situación, la clínica va a aparecer sobre todo en la etapa de transición y va a depender de diversos factores, entre ellos, de las coinfecciones con otros patógenos, especialmente infecciones bacterianas secundarias y la presencia del virus del síndrome reproductivo y respiratorio porcino (PRRSV). En diversos estudios se estima que el impacto de la gripe enzoótica puede llegar a aumentar hasta un 2% la mortalidad en la etapa de transición, así como las infecciones bacterianas secundarias y los costes médicos derivados (Torremorell et al., 2009; Gillespie et al., 1999). También se ha visto que la infección temprana durante la lactancia por este virus puede aumentar la mortalidad después del destete, lo cual sugiere una posible interacción con otros patógenos (Álvarez et al., 2015).

Para comprender estas sinergias hay que entender cómo es la patogenia de este virus y cómo ella facilita o agrava la infección de otros patógenos. Por otro lado, para focalizar el control es importante evaluar qué patógenos van a resultar más importantes en el agravamiento del cuadro clínico y en el impacto en parámetros productivos. En este artículo se resume brevemente de qué conocimiento se dispone actualmente en estos aspectos.
La patogenia del VIA: sinergias con bacterias
En los mamíferos, como en el cerdo, el VIA causa una infección aguda que se produce después de la inhalación de aerosoles que contienen partículas víricas (ver figura). La infección va a estar limitada al tracto respiratorio, aunque el virus se puede detectar en otros tejidos, como los ganglios linfáticos locales, pero sin replicarse (Kuiken et al., 2012). A las pocas horas después de su inhalación, se puede detectar en el núcleo y citoplasma de células epiteliales (células ciliadas y células secretoras) de la tráquea y bronquios (Kuiken et al., 2012). Durante las primeras 24 h se observa cómo los viriones empiezan a salir al lisarse de las células infectadas y avanzan en la infección de nuevas células hasta bronquiolos y alvéolos. La necrosis de las células ciliadas secretoras causada por la infección del VIA va a limitar la capacidad del pulmón de expulsar bacterias, lo que, a su vez, causa un aumento de la concentración bacteriana en el tracto respiratorio. Por tanto, ésta sería una primera acción sinérgica entre el VIA y otros patógenos.
Por otro lado, para que los viriones se liberen de las células, el virus debe “desengancharse” del receptor celular, algo que va a conseguir mediante la acción de la neuraminidasa (N). Esta glicoproteína de superficie va a cortar el ácido siálico presente en los receptores celulares y, con ello, va a aumentar la afinidad del receptor con bacterias que antes no se podían adherir en la superficie de las células (McCullers et al., 2016). Ésta sería una segunda interacción entre el virus de la gripe y las bacterias. En tercer lugar, debido a la replicación y liberación del VIA se produce necrosis celular y descamación de restos celulares que pueden llegar hasta la luz alveolar. Recordar que la concentración de las bacterias también es mayor y muchas están adheridas a la superficie de estos restos celulares, lo que puede servir para “sembrar” otras partes del pulmón.
Finalmente, el virus de la gripe puede causar, por razones todavía no bien estudiadas, la depleción de macrófagos alveolares, lo cual diezma una de las primeras barreras de la inmunidad innata. Esta depleción dura alrededor de dos semanas y abre una ventana que puede permitir la entrada de otros patógenos.

Coinfecciones del VIA con otros patógenos en la especie porcina
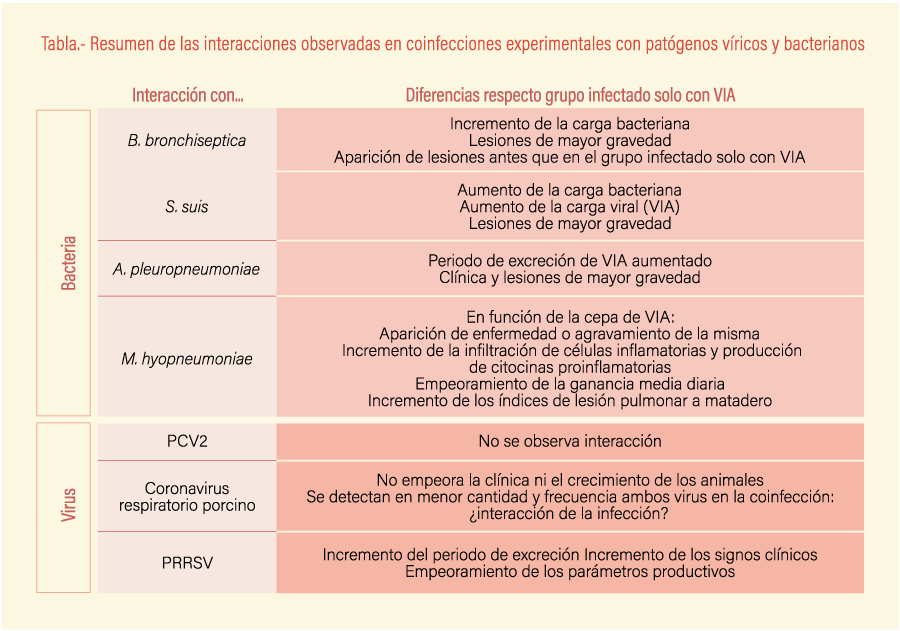
Existen diferentes estudios en los que se valora la coinfección del virus de la gripe con otros patógenos (ver tabla). En muchos de ellos, se observa un incremento de la gravedad de las lesiones de ambos patógenos por separado, así como un empeoramiento del cuadro clínico. Esto es especialmente cierto cuando se producen coinfecciones del VIA con bacterias, pero no es tan evidente con las coinfecciones con otros virus.
Interacción del VIA con bacterias
Como se ha resumido en el apartado anterior, la patogenia del virus de la gripe genera un ambiente en el tracto respiratorio que facilita claramente la aparición o agravamiento de infecciones bacterianas secundarias.
Existen diferentes infecciones experimentales publicadas que demuestran que esta interacción es real con bacterias importantes en la etapa de transición como Bordetella bronchiseptica o Streptococcus suis. Con la primera de ellas se observa un incremento de la colonización de esta bacteria y la aparición de lesiones (Loving et al., 2010). Además, en este estudio se observó que la combinación VIA-B. bronchiseptica aceleraba la aparición de lesiones respecto al grupo de animales infectados únicamente por el VIA, siendo además de mayor gravedad y aumentando la afectación pulmonar. Estos resultados son parecidos en el caso del Streptococcus suis; la concentración de unidades formadoras de colonias de S. suis en animales coinfectados con el VIA es significativamente superior a animales infectados únicamente con la bacteria. De nuevo, las lesiones son de mayor gravedad (Lin et al., 2015) para la coinfección. En este caso, además, la concentración del virus también se ve aumentada tanto en hisopo nasal como en pulmón, lo cual indica claramente la sinergia entre ambos agentes.
¿Qué sucede en la fase de cebo?
En cuanto a bacterias de importancia en el engorde, se describen sinergias con Actinobacillus pleuropneumoniae y con Mycoplasma hyopneumoniae. En infecciones experimentales se observa que, en animales coinfectados, el periodo de excreción vírica se ve aumentado y la clínica y las lesiones son más graves (Pomorska-Mol et al., 2017). El caso de M. hyopneumoniae es interesante ya que causa la misma lesión que el VIA (neumonía bronquiolointersticial) aunque el cuadro clínico suele aparecer en momentos distintos.
Pero, ¿qué sucede cuando la infección se produce por ambos patógenos? En dos estudios realizados por Deblanc et al. (2012 y 2016) se demostró que la gravedad de la infección del virus de la VIA era distinta en función del subtipo de gripe y de la preinfección por M. hyopneumoniae. En este estudio se usaron dos VIA distintos: un H1N1 y un H1N2. Se observó que el H1N1 por sí solo no era capaz de provocar sintomatología clínica ni lesiones graves, mientras que el H1N2, sí. En cambio, cuando la infección por los VIA se realizaba después de una primoinfección por M. hyopneumoniae, el H1N1 provocaba clínica y lesiones de la misma gravedad que el H1N2, mientras el grupo infectado por este segundo virus no presentaba cambios significativos con respecto al grupo de animales infectados solamente por H1N2. Además, la ganancia media diaria se veía significativamente reducida en los grupos coinfectados.
Posteriormente, se observó que M. hyopneumoniae predispone a una mayor inflamación con una mayor infiltración de neutrófilos y macrófagos, así como un claro incremento de la producción de citocinas proinflamatorias, lo que explicaría el cambio en la gravedad observada para el virus H1N1.
Finalmente, es remarcable que las explotaciones seropositivas a distintos subtipos de gripe tienden a presentar peores índices de lesión pulmonar en matadero (Fablet et al., 2018).
Interacción del VIA con otros virus
Existen estudios de coinfección en condiciones experimentales entre el VIA y el coronavirus respiratorio porcino (PRCoV, el circovirus porcino tipo 2 (PCV2) y con el PRRSV. El primer estudio (Lanza et al., 1992) muestra que la combinación de H1N1 o H3N2 con PRCoV no aumenta la gravedad del cuadro clínico ni de las lesiones. Tampoco se observa un impacto en el crecimiento de los animales en animales coinfectados si se compara con animales infectados con uno solo de los virus. Interesantemente, ambos virus se detectaron con menor frecuencia en animales coinfectados, lo que sugiere una posible interferencia entre ambos patógenos. En el caso del PCV2, tampoco se observa un efecto sinérgico con el VIA en cuanto a clínica, carga vírica o lesiones (Wei et al., 2010), aunque sí se ha asociado su presencia en explotaciones porcinas (Jiménez et al., 2014).
Un caso aparte es la coinfección con el virus del PRRSV. En un estudio realizado por Van Reeth et al. (1996) se observó que la infección de VIA después de haber infectado con PRRSV aumentaba el periodo de excreción del VIA hasta dos días más del periodo normal. Además, en parte de los animales coinfectados se observó un aumento significativo de la temperatura corporal y de los signos clínicos comparado con los animales infectados con un solo virus, además de retrasar el crecimiento de los animales coinfectados. Es complicado valorar si las explotaciones que presentan ambos patógenos sufren un empeoramiento de los parámetros productivos o sanitarios, probablemente debido a la elevada prevalencia de ambos y la dificultad de encontrar explotaciones infectadas con uno de ellos pero, si se tiene en cuenta los resultados de este estudio, es esperable que haya algún tipo de impacto.
Conclusión
El VIA es un agente claramente primario que actúa como agente primario en infecciones bacterianas y agrava su cuadro clínico. Tiene impacto a nivel económico y productivo en las explotaciones de porcino, sobre todo en su forma enzoótica. En algunos casos (por ejemplo, en combinación con PRRSV o M. hyopneumoniae), incluso se podría considerar como un agente secundario, agravante o, como mínimo, sinérgico.
Actualmente, se sabe que tanto el viroma como la microbiota nasal del cerdo son muy complejos, lo que abre muchas posibilidades en el estudio de asociaciones entre el VIA y otros virus y/o bacterias en un futuro próximo.
Bibliografía:
Torremorell, et al. (2009). Vet Rec. 165(3):74-77. Gillespie, et al. (1999). Swine Health Prod. 7(2):81–83.
Alvarez, et al. (2015). Prev Vet Med. 121(3-4):240-245.
Kuiken, et al. (2012). Current Op in Virol, 2:276–286.
McCullers (2016). Nat Rev Micr. 12:252-262.
Loving, et al. (2010). Microb Pathog. 49(5):237-245.
Lin, et al. (2015). PLoS One. 10(4):e0124086.
Pomorska-Mol, et al. (2012). FEMS Immunol Med Microbiol. 2012 66(3):334-342.
Deblanc, et al. (2012). Vet Microbiol. 157(1-2):96-105.
Deblanc, et al. (2016). J Gen Virol. 97(10):2501-2515.
Fablet, et al. (2018). Porcine Health Management 4:6. Lanza, et al. (1992). Res Vet Sci. 53(3):309-14.
Wei, et al. (2010). Comp Med. 60(1):45-50.
Jimenez, et al. (2014). Virol Sin. 29(4):242-9.
Van Reeth, et al. (1996). Vet Microbiol. 48(3-4):325- 35.