
Interacciones del virus de la gripe con otros patógenos
22 abril, 2021
Inmunología y vacunas: respuesta a las vacunas y planes vacunales frente a la gripe porcina
29 abril, 2021Artículo publicado en Suis nº 165 - Marzo 2020
Carlos Casanovas, Salvador Oliver, Sonia Cárceles, Laura Garza y David Espigares
Servicio Técnico Porcino CEVA Salud Animal
Introducción
Como en la especie humana, la especie porcina se ve afectada por virus de la Influenza del tipo A (AIV), compartiendo los subtipos H1N1 y H3N2 (el H1N2 afecta a la especie porcina, pero no a la humana). Además, ambas especies pueden infectarse por el linaje pandémico del H1N1 (H1panN1), una variante del H1N1 que en el año 2009 generó mucha alarma al producir una pandemia en la especie humana. Este H1panN1 se originó en la especie porcina tras múltiples reordenaciones con virus aviares, humanos y porcinos y acabó infectando a la especie humana que fue la responsable de su transmisión a nivel mundial. Hoy en día, el H1panN1 se encuentra en ambas especies a nivel mundial.
El hecho de que ambas especies se vean afectadas por los mismos AIV no es casualidad. La infección se inicia mediante la unión de la hemaglutinina (HA) de la superficie del virus con los azúcares de ácido siálico de la superficie de las células epiteliales respiratorias y ambas especies comparten los mismos receptores de tipo α2-6. Además, la estructura anatómica de los órganos internos es también muy parecida. Esto hace que ambas especies puedan infectarse entre ellas con cierta facilidad y compartir AIV del mismo tipo.
Estas similitudes entre ambas especies nos permiten extrapolar investigaciones realizadas en la especie humana a la especie porcina.
La patogenia del AIV
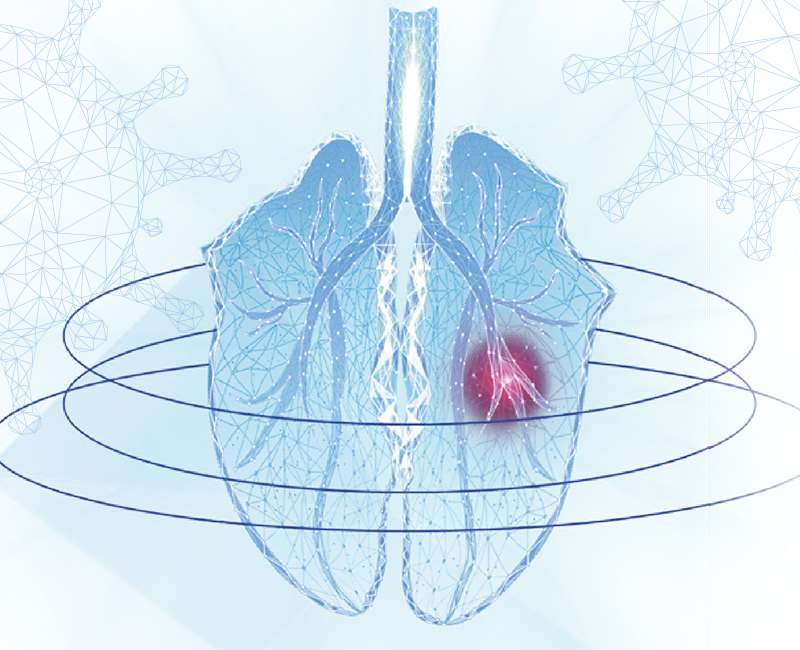
AIV accede por vía nasofaríngea y se une a células epiteliales de la mucosa nasal, etmoides, tráquea, bronquios y bronquiolos pulmonares, donde empieza a replicar, causando la degeneración y muerte de las células que se desprenden hacia el lumen (Figura 1). En poco tiempo se genera respuesta inflamatoria con abundantes células inflamatorias (neutrófilos en mayor parte) y producción de citoquinas. La obstrucción causada por las células necróticas y el exudado inflamatorio colapsa las vías respiratorias menores, bloquea el flujo de aire y generan una consolidación pulmonar difusa con aparición en 24-48 horas de lesiones microscópicas (neumonía broncointersticial) características de AIV.
A los 5-7 días, el virus cesa su replicación, desaparecen las células necróticas y la inflamación y los tejidos empiezan a regenerarse, por lo que, en ausencia de complicaciones secundarias, se recupera la normalidad alrededor de los 14-21 días tras la infección (Figura 1).
Sin embargo, en el ámbito de una explotación porcina, con presencia de múltiples agentes infecciosos tanto víricos como bacterianos, las complicaciones secundarias favorecidas por las lesiones provocadas por AIV suelen ser frecuentes.

Figura 1. Patogenia de AIV
Mecanismos que favorecen las complicaciones bacterianas secundarias
Obstrucción de vías respiratorias y alteración del movimiento ciliar
La propia obstrucción de vías respiratorias con presencia de células respiratorias y exudado inflamatorio genera espacios muertos que representan un caldo de cultivo ideal para la proliferación de bacterias. A esto se le añade el compromiso del movimiento ciliar que dificultará el despeje de bacterias.
Alteración del surfactante
Los neumocitos alveolares de tipo 2 son los responsables de la producción de surfactante, que tiene la doble función de evitar el colapso alveolar y también actúa de barrera impidiendo que los agentes infecciosos sean capaces de atravesar los capilares y llegar al torrente sanguíneo. La destrucción de neumocitos causada por AIV contribuye al colapso del pulmón y al paso de agentes como Streptococcus suis a sangre, que luego puede tener su lugar de acción en sitios tan diversos como las meninges o las articulaciones.
Incremento de la exposición de receptores frente a bacterias
La necrosis de células epiteliales causada por AIV, expone lugares de adhesión para las bacterias y la respuesta inflamatoria altera la regulación y exposición de proteínas en la superficie del epitelio respiratorio. Los cambios durante la fase de recuperación de las lesiones también generan lugares de adhesión. En zonas con curación incompleta se exponen elementos de la membrana basal (laminina, colágeno) o depósitos de fibrina y fibrinógeno que favorecen la unión de las adhesinas bacterianas. Por este motivo, en algunos casos, se observa que la complicación bacteriana ocurre cuando empieza a haber recuperación de la infección primaria.
Tropismo y características de la Hemaglutinina de AIV
En la especie humana se ha visto que los AIV con preferencia por uniones a receptores α2,3 generan infecciones pulmonares más profundas, a nivel de vías respiratorias bajas. También se sabe que los AIV con Hemaglutininas con alto nivel de glicosilación se unen con mayor facilidad al surfactante, por lo que el virus es eliminado con mayor facilidad por los cilios. Curiosamente, recientemente se ha descubierto que el AIV que generó la pandemia de 1918 (Gripe Española) tenía capacidad para unirse a receptores α2,3 y además tenía una HA con poca glicosilación, lo que predispuso a la aparición de neumonías bacterianas secundarias. Esta pandemia mató entre 1918 y 1920 a más de 40 millones de personas y se considera la más grave de la historia. La mayoría de muertes fueron debidas a neumonías bacterianas secundarias, ya que en esa época no había antibióticos disponibles
Como vemos, en la especie humana no todas las cepas de AIV muestran el mismo nivel de patogenicidad y esto es también extrapolable a la especie porcina. El hecho de que la cepa tenga la capacidad de colonizar las vías respiratorias bajas (pulmón) es determinante en su patogenicidad. Este hecho, sumado a las características de su HA pueden jugar un papel determinante a la hora de predisponer infecciones bacterianas secundarias
Efectos del virus en la inmunidad frente a bacterias
En la especie humana se ha visto que la NS1, una proteína interna no estructural de AIV interfiere en la inmunidad pulmonar innata frente a las bacterias. Además, AIV consume los macrófagos alveolares responsables del bloqueo de las infecciones bacterianas tempranas y se necesitan más de 2 semanas para reemplazarlos. En presencia de esta actividad fagocitaria deprimida, la inflamación mediada por neutrófilos causaría lesiones sin lograr un control efectivo de patógenos.
Además, durante la fase de limpieza de AIV y de curación de lesiones se establece un estado antiinflamatorio general regulado por el aumento de IL-10. Esta citoquina suprime los mecanismos involucrados en el reconocimiento y limpieza de patógenos, para lograr la recuperación de la homeostasis pulmonar. En esta situación también hay una depresión de la capacidad fagocítica de macrófagos y neutrófilos, que favorecería las complicaciones por otros agentes infecciosos. Este hecho también contribuiría a que, en algunas ocasiones, la complicación bacteriana ocurriera cuando empezara a haber recuperación de la infección primaria.
Aumento de la Inflamación
En la especie humana, también se ha descrito que algunos tipos de PB1 (una proteína interna de AIV) causan mayor citotoxicidad y activación de la respuesta inflamatoria, por lo que, de nuevo, algunas cepas de AIV serían más capaces de inducir inflamación que otras. Además, las bacterias también promueven la producción de citoquinas, lo que aumentaría la sinergia con AIV.

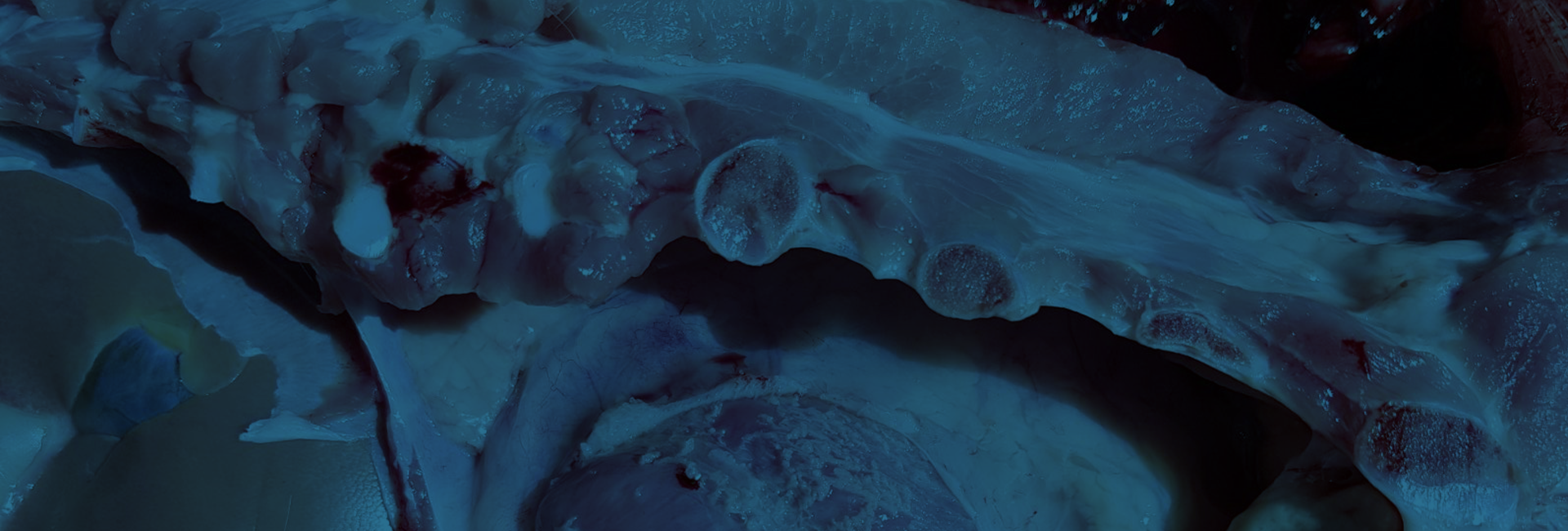
Figura 2. Poliserositis
Principales agentes bacterianos secundarios que afectan al porcino
Hasta ahora hemos visto distintos mecanismos mediante los cuales AIV abre la puerta a la colonización del pulmón por bacterias. Entre estas, las principales son: Bordetella bronchiseptica, Haemophilus parasuis, Streptococcus suis, Mycoplasma hyorhinis, Pasteurella multocida, Mycoplasma hyopneumoniae y Actinobacillus pleuropneumoniae
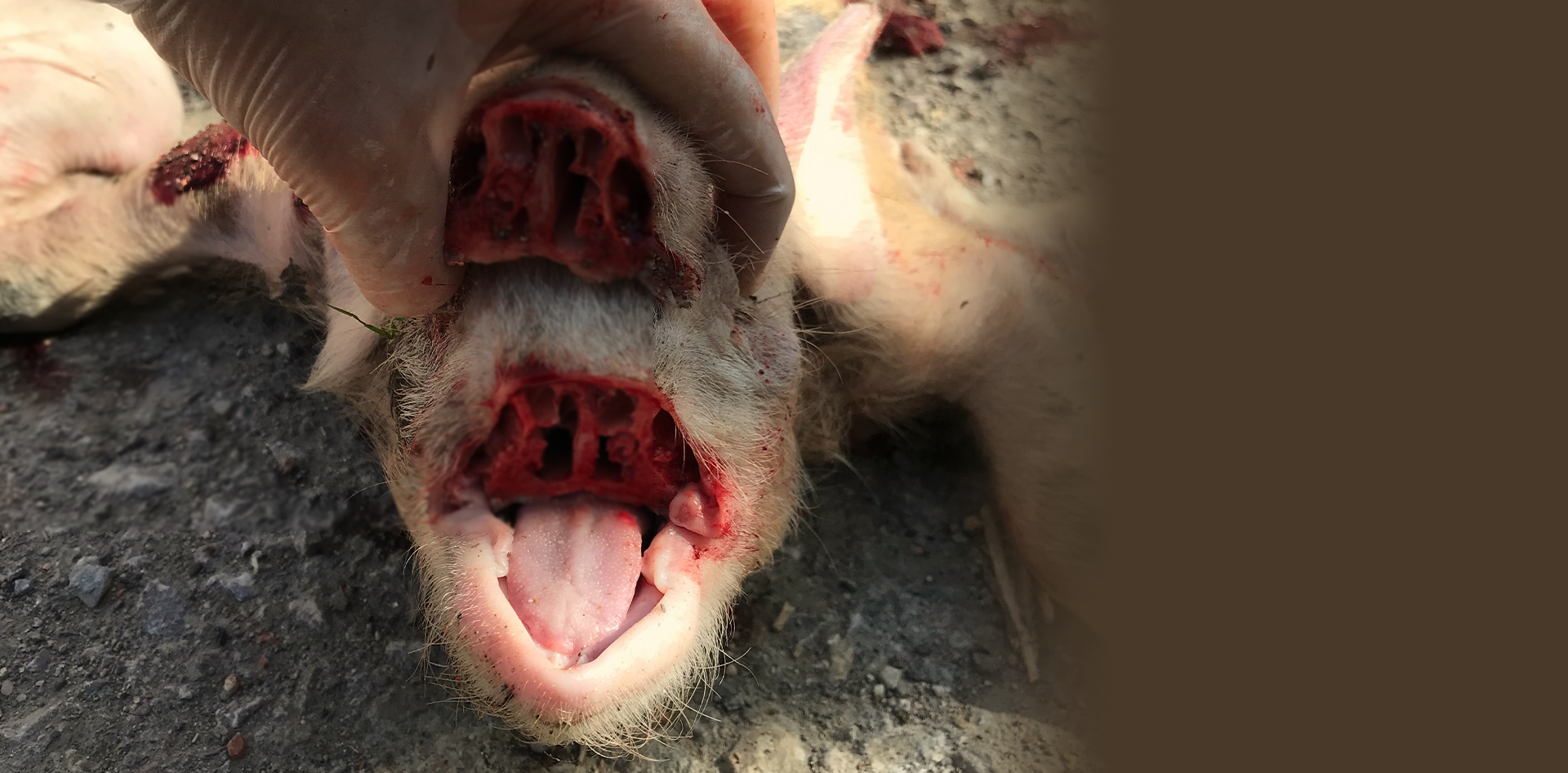
Hoy en día, en la fase de transición se describe un aumento importante de los casos de poliserositis (Figura 2), artritis y meningitis, tras la retirada de medicaciones (fundamentalmente amoxicilina) del pienso. Agentes como H. parasuis y S. suis, se encuentran con frecuencia en la cavidad nasal y a menudo necesitan la colaboración de agentes primarios como PRRSV o AIV para acceder al pulmón. Otro factor importante en este proceso es la participación de agentes infecciosos y/o situaciones ambientales que favorezcan rinitis atrófica. Un elevado porcentaje de lechones en fase de transición de muchas explotaciones sufren problemas de rinitis atrófica, probablemente no progresiva, porque solo cuentan con la participación de B. bronchiseptica (sin la participación de cepas toxigénicas de P. multocida tipo D y A). En esta situación, a las lesiones descritas en el epitelio respiratorio causadas por AIV, se le suma la destrucción de cornetes nasales (Figura 3), lo que permite un acceso directo al pulmón de los agentes que se encuentran en cavidad nasal.

Figura 3. Destrucción de los cornetes nasales
Conclusiones
En este artículo hemos visto que AIV es un patógeno primario cuyo grado de patogenicidad difiere entre cepas, y que en la especie humana ya se conocen algunos de los factores que afectan a la virulencia de las cepas. Estos factores son probablemente extrapolables a la especie porcina ya que ambas especies guardan elevado grado de similitud y los virus que las afectan también. Por otro lado, las alteraciones que provoca AIV a nivel de vías respiratorias promueven la infección por bacterias secundarias presentes de forma habitual en la mayoría de explotaciones, más aún tras la disminución del uso de antibióticos. Esto significa que, en la situación actual de disminución del uso de antibióticos, el control del complejo respiratorio porcino pasa por controlar agentes primarios como PRRSV y AIV, los cuales agravan infecciones bacterianas secundarias. Dentro de este complejo también es clave controlar factores que inducen la presencia de rinitis.
Bibliografía:
Casas, Josep et al. OvisLab S.L. ¿Son un Complejo las poliserositis? revista Suis nº 124, enero-febrero 2016.
Kuiken T et al. Pathogenesis of influenza virus infections: the good, the bad and the ugly. Current Opinion in Virology 2012, 2:276–286.
McCullers Jonathan A. The co pathogenesis of influenza viruses with bacteria in the lung. NATURE abril 2014 vol. 12.
Zimmerman, Jeffrey J. Diseases of swine 10th edition, capítulo 40, Influenza viruses. Wiley-Blackwell.